
Fizjologia słyszenia - czyli jak to się dzieje, że słyszymy?
Fale akustyczne, percepowane w narządzie słuchu jako dźwięk, wprawiają w drgania mechaniczne układ transmisyjny ucha środkowego. Zakres częstotliwości słyszalnych przez człowieka obejmuje częstotliwości od 16 Hz do około 20 000 Hz. Do narządu Cortiego dźwięk dociera na drodze przewodnictwa powietrznego i kostnego. Energia akustyczna zostaje przetworzona w uchu środkowym w energię mechaniczną, która w dalszych odcinkach narządu słuchu zamieniana jest w energię bioelektryczną.
Fale dźwiękowe zbierane są przez małżowinę uszną, a następnie skierowane do przewodu słuchowego. Następuje drganie błony bębenkowej. Drgania zostają przekazane na kosteczki słuchowe: młoteczek, kowadełko i strzemiączko. Strzemiączko przenosi drgania na błonę okienka owalnego ślimaka, a ta z kolei na perylimfę w schodach przedsionka. Ruchy płynu przedsionka przekazywane są do płynu w przewodzie ślimakowym i przez błonę podstawną do płynu schodów bębenka. Między schodami przedsionka i schodami bębenka leżą schody środkowe w których leży narząd Cortiego. Odkształcanie błony okienka okrągłego wygasza wibracje perylimfy, co umożliwia przyjmowanie i rejestrowanie kolejnych dźwięków. Drgania błony podstawnej wywołane drganiami perylimfy zostają przeniesione na narząd Cortiego. Narząd Cortiego (spiralny) zawiera receptory słuchu. Zbudowany jest z komórek podporowych i włoskowych (około 16 000) ułożonych w szeregi. Na wolnej powierzchni komórek włoskowych znajdują się stereocylia. Do komórek włoskowych docierają neuryty dośrodkowe (akustyczne i spiralne), neuryty odśrodkowe (od jądra oliwki) i neuryty współczulne.
W skład narządu Cortiego wchodzą dwa rodzaje komórek słuchowych: ułożone w pojedynczym rzędzie komórki słuchowe wewnętrzne w liczbie ok. 3,5 tys. oraz ułożone w trzech rzędach komórki słuchowe zewnętrzne w liczbie ok. 12 tys. Komórki słuchowe są wyspecjalizowane w odbieraniu bodźców o rożnych częstotliwościach. Komórki odpowiedzialne za słyszenie wysokich częstotliwości znajdują się przy podstawie ślimaka, a im dalej od strzemiączka, tym niższe częstotliwości odbierane są przez komórki słuchowe. Oba typy komórek posiadają w części szczytowej rzęski. Komórki słuchowe wraz z komórkami podporowymi znajdują się na błonie podstawnej narządu Cortiego. Ich górna, zakończona rzęskami część komunikuje się z galaretowatą strukturą zwaną błoną nakrywkową. W przypadku komórek słuchowych zewnętrznych ich rzęski zatopione są w błonie nakrywkowej. Rzęski komórek słuchowych wewnętrznych nie są scalone z błoną nakrywkową, lecz znajdują się w jej bezpośredniej bliskości. Komórki słuchowe unerwione są przez 2 rodzaje włókien nerwowych: włókna aferentne (przewodzące bodźce dośrodkowo, tzn. od ślimaka do ośrodkowego układu nerwowego), czyli włókna nerwu słuchowego, w liczbie ok. 30 tys. biegnące z obwodu do ośrodków centralnych, oraz włókna eferentne (odśrodkowe, czyli przewodzące bodźce od ośrodkowego układu nerwowego do ślimaka), zwane oliwkowo-ślimakowymi w liczbie kilkuset, biegnące od oliwki na obwód do ślimaka. Zdecydowana większość (93%) włókien aferentnych ślimaka unerwia komórki słuchowe wewnętrzne, jedynie 7% włókien aferentnych związanych jest z komórkami słuchowymi zewnętrznymi. Niemal wyłącznym unerwieniem komórek słuchowych zewnętrznych są włókna eferentne.
Włókna aferentne przewodzą pobudzenie do ośrodkowego układu nerwowego i pozwalają na powstanie wrażenia dźwięku. Włókna eferentne, według badań, pełnią rolę ochronną w urazie akustycznym, hamując nadmierną odpowiedź ślimaka w przypadku zbyt silnych bodźców, które mogłyby uszkodzić narząd Cortiego. Pobudzenie włókien eferentnych ślimaka występuje na zasadzie sprzężenia zwrotnego po pobudzeniu włókien aferentnych i dzięki istnieniu włókien skrzyżowanych jest ono obuuszne. Komórki słuchowe zewnętrzne są bardzo wrażliwe na działanie czynników uszkadzających i ulegają uszkodzeniu w pierwszej kolejności. Wybiórcze zniszczenie komórek słuchowych zewnętrznych powoduje uszkodzenie słuchu rzędu 30-50 dB.
Zniszczenie komórek słuchowych wewnętrznych powoduje całkowitą głuchotę.
Drgania błony podstawnej na której leży narząd Cortiego powoduje zginanie stereocylii. Otwarte zostają wówczas kanały jonowe dla potasu. Potas przenika do cytozolu stereocyliów co wywołuje depolaryzację błony komórek włoskowych i wytworzenie potencjału czynnościowego. Lokalizacja odgięcia stereocylii w komórkach włoskowych decyduje o wysokości dźwięku. Tony wysokie powstają przy okienkach, średnie po środku ślimaka, a niskie w tzw. szczycie ślimaka. Amplituda odgięć stereocylii determinuje natężenie dźwięku. Podniety początkowo biegną do zwoju spiralnego ślimaka, od którego odchodzi nerw ślimakowy. Nerw ślimakowy biegnie do jąder ślimakowych leżących w trzonie mózgu.
W jądrach następuje przekazanie impulsów do neuronów czuciowych. Impulsy biegną do mostu (ciało czworoboczne). W tak zwanym jądrze górnym oliwki mieści się początek drogi słuchowej biegnącej do wzgórków czworaczych. Pomiędzy lewym i prawym wzgórkiem istnieje połączenie, co umożliwia neurotransmisję impulsów na przeciwną stronę ciała. Od wzgórków czworaczych neuryty biegną do ciała kolankowatych (lewy i prawy), a od nich do ośrodków słuchowych w korze mózgowej (płaty skroniowe). W ciałach kolankowatych przyśrodkowych następuje integracja informacji akustycznych.
W korze mózgowej zachodzi projekcja i kojarzenie informacji akustycznych.
Narząd równowagi jest źródłem informacji o pozycji ciała. Dzięki temu możliwy jest odpowiedni rozkład napięcia w różnych grupach mięśniowych. Zbudowany jest z trzech kanałów półkolistych: przedniego, tylnego i bocznego, ustawionych prostopadle do siebie i uchodzących do łagiewki. Pomiędzy łagiewka i ślimakiem znajduje się woreczek. W ścianie woreczka i łagiewki znajdują się plamki statyczne, pokryte nabłonkiem walcowatym, w którym mieszczą się mechanoreceptory. Na powierzchni mechanoreceptorów znajdują się krótkie stereocylia i 1 długie kinetocylium. Na powierzchni plamek statycznych leży warstewka substancji żelowatej na której znajdują się wapienne kamyczki błędnikowe.
Kanały półkoliste wypełnione są endolimfą. Ujścia kanałów są rozszerzone, tworząc bańki. Wysłane są nabłonkiem płaskim. Niektóre obszary – grzebienie baniek wysłane są nabłonkiem walcowatym zawierającym komórki włosowate. Komórki włosowate pokryte są galaretowatym osklepkiem na którym leż kamyczki wapienne – otolity. Endolimfa zawiera dużo jonów potasu. Zmiany położenia ciała powodują ruch endolimfy oddziałujący na kamyczki – otolity. Naciskając na stereocylia, powodują ich odgięcie w kierunku kinetocylium. Powoduje to otwarcie kanałów jonowych (białka śródbłonowe) dla potasu, który przenika do cytozolu stereocyliów. Efektem tego jest depolaryzacja błony, czyli pobudzenie komórek zmysłowych. Odgięcie stereocyliów w drugim kierunku – od kinetocylium powoduje hiperpolaryzację i zahamowanie przewodzenia. Zwój przedsionkowy zbiera neuryty od komórek zmysłowych. Od zwoju odchodzi nerw przedsionkowy, biegnący do mostu (do jądra przedsionkowego). Z jąder przedsionkowych impulsy przekazywane są do rdzenia, móżdżku oraz kory mózgowej. Na drodze odruchowej następuje wzmożenie napięcia określonych grup mięśni, co przywraca równowagę.
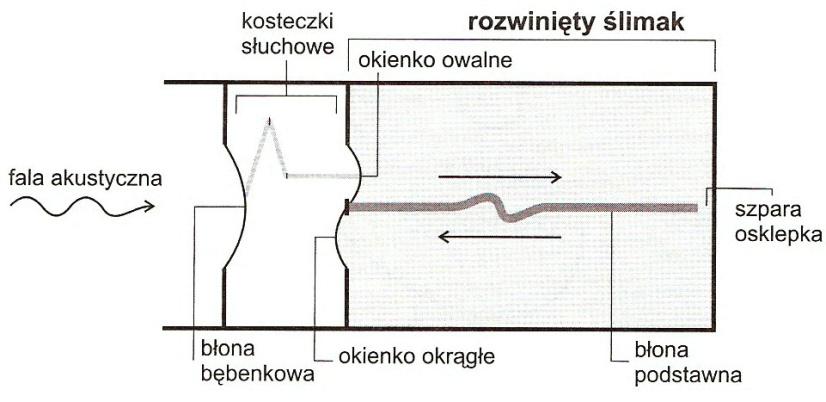
Specyficzna budowa błony bębenkowej związana jest z pełnym przekazywaniem częstotliwości istotnych dla rozumienia mowy ludzkiej (częstotliwości średnie). Oznacza to, że błona bębenkowa dla tych częstotliwości przyjmuje pełną energię fali akustycznej, a tylko mała część fal jest odbijana (opór – impedancja – jest niewielki). Zwiększona oporność (impedancja) błony bębenkowej występuje w przypadku, gdy ciśnienie w jamie bębenkowej jest niższe od ciśnienia w przewodzie słuchowym zewnętrznym. Zmiana impedancji błony bębenkowej związana jest też ze skurczem m. strzemiączkowego i m. napinającego błonę bębenkową. Pomiary impedancji ucha środkowego są bardzo istotnymi badaniami w diagnostyce uszkodzeń narządu słuchu. Drgania błony bębenkowej przenoszone są przez kosteczki słuchowe do przestrzeni przychłonkowej ucha wewnętrznego.
Różnica pomiędzy wielkością powierzchni błony bębenkowej, a płytki strzemiączka oraz przekładnia dźwigniowa kosteczek słuchowych zmniejszają amplitudę drgań i wzmacniają działanie układu transmisyjnego ucha środkowego w stosunku od około 1 : 18 do 1 : 22. Opisane mechanizmy stwarzają korzystne warunki dla fali dźwiękowej przechodzącej ze środowiska powietrznego (niska impedancja) do środowiska płynnego (wysoka impedancja) ucha wewnętrznego. Mięśnie ucha środkowego tłumią drgania łańcucha kosteczek po zadziałaniu bodźca. Pełnią one również funkcję ochronną w odniesieniu do ucha wewnętrznego, ponieważ przy skurczu usztywniają łańcuch kosteczek słuchowych, ograniczając w ten sposób dopływ energii (mechanizm ochronny wyzwalany przy dźwięku o znacznym natężeniu).
Przewodnictwo kostne można zbadać przy użyciu stroików (kamertonów) lub stosując do badania słuchawkę dla przewodnictwa kostnego. Przewodzenie dźwięku na drodze przewodnictwa kostnego odbywa się poprzez bezpośrednie wprawianie w drgania przychłonki (perilympha) przenoszone przez kości czaszki, jak również przez ucho środkowe.
W uchu wewnętrznym dokonywana jest analiza dźwięku. Według teorii rezonansowej Helmholtza błona podstawna ślimaka zbudowana jest z włókien poprzecznych o różnej długości i różnym napięciu. Włókna te działają jako rezonatory i w drgania wprawiane zostają tylko te włókna, których częstotliwość drgań jest zgodna z częstotliwością fali dźwiękowej. Teoria ta ma obecnie już tylko znaczenie historyczne. Aktualnie powszechnie uznawana jest teoria hydrodynamiczna von Bekesyego i Rankego, oparta na bezpośrednich obserwacjach modeli ślimaka i preparatach anatomicznych. Została ona również udokumentowana w badaniach eksperymentalnych i teoretycznych. Zgodnie z hydrodynamiczną teorią ruch płytki strzemiączka wywołuje zmianę objętości graniczącej z nim przychłonki (warunkiem jest ruchoma płytka strzemiączka). Zmiana objętości przychłonki wychyla błonę podstawna wraz z całym przewodem ślimaka najpierw na jego początku, a potem wychylenia przenoszone są jako fala wędrująca z różną prędkością i zasięgiem od płytki strzemiączka, do osklepka.
Zwiększająca się szerokość błony podstawnej, jej elastyczność oraz zmniejszający się przekrój kanału kostnego nadają fali wędrującej szczególny charakter. Amplituda fali wzrasta do miejsca maksymalnego wychylenia i załamuje się, podobnie do fali na płaskiej plaży. W tym punkcie dochodzi do dyspersji, czyli przestrzennego podzielenia dźwięku według częstotliwości. Drgania dla wysokich częstotliwości mają swoje maksymalne amplitudy w pobliżu płytki strzemiączka, a fale dla częstotliwości niskich w pobliżu szpary osklepka.
Tak więc każda częstotliwość przy porządkowana jest do określonego miejsca wychylenia, jak to już wcześniej przedstawił Helmholtz (teoria jednego miejsca, tonotopowa organizacja ślimaka) z tą jednak różnicą, że przez dyspersję, a nie przez rezonans.
Przy wychyleniu błony podstawnej i przesunięciu błony pokrywającej (membrana tectoria) lub śródchłonki działają siły nożycowe, które przesuwają włoski czuciowe i przedstawiają adekwatny bodziec.
Mechanizm pobudzenia komórki zmysłowej
Transformacja dźwięku
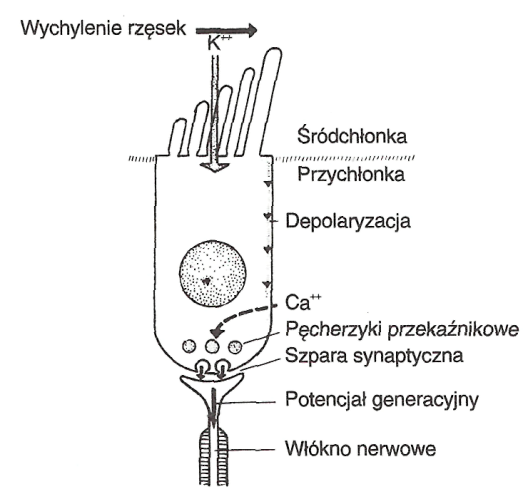
W ślimaku energia mechaniczna zostaje przekształcona w energię elektryczną. Pojedyncze bodźce nerwowe w komórkach zmysłowych wywołane są w wyniku ich pobudzenia przez akustyczny sygnał powstały w miejscu maksymalnej amplitudy. Wówczas w komórkach zmysłowych występuje synchroniczna zmiana potencjału receptorowego, który przez potencjał generacyjny po przekroczeniu określonego progu (zasada wszystko albo nic) wywołuje potencjał czynnościowy we włóknach nerwowych. Przy ruchu rzęsek komórek zmysłowych, związanym z kierunkiem wychylenia błony, następuje otwarcie kanałów jonowych w części wierzchołkowej błony komórek zmysłowych, co prowadzi do dostarczenia jonów potasu ze śródchłonki. Zjawisko to wywołuje depolaryzację błony komórki zmysłowej i związany z tym przepływ jonów wapnia z cortilymfy. Reakcje te prowadzą do uwolnienia zawartości z pęcherzyków przekaźnikowych w szparze synaptycznej i powstania postsynaptycznego potencjału generacyjnego. Repolaryzacja następuje poprzez powrotny transport jonów, co wymaga energii. Zmiany napięcia potencjałów w ślimaku mogą być rejestrowane na promontorium jako potencjały mikrofoniczne ślimaka (cochler microphonis). Bodźce zmysłowe są przekazywane przez komórki włosowate wewnętrzne. Komórki zmysłowe zewnętrzne, obok zdolności przemiany energii akustycznej w energię elektryczną (transdukcja mechaniczno-elektryczna), posiadają również zdolności motoryczne związane z wyposażeniem ich w aktynę. W wyniku tego aktywnego procesu dochodzi do wzmocnienia amplitudy fali wędrującej i tłumienia wychyleń sąsiednich odcinków błony podstawnej. Ten ślimakowy przedwzmacniacz umożliwia komórkom włosowatym wewnętrznym reakcję również na słabe bodźce akustyczne, co wyrażone jest zwiększoną wrażliwością słuchu. W zależności od bodźca akustycznego układ eferentny reguluje czynności komórek zmysłowych zewnętrznych. Komórki zmysłowe zewnętrzne mają zdolność do kurczenia się i emitowania odbitej od nich części fali dźwiękowej docierającej do ucha wewnętrznego, co tworzy podstawę dla emisji otoakustycznej. Zjawisko to związane jest z ruchem przychłonki w odwrotnym kierunku, poprzez ruch kosteczek słuchowych i błony bębenkowej; otoemisję w postaci sygnałów akustycznych ucha wewnętrznego można zmierzyć w przewodzie słuchowym zewnętrznym.
Wytwarzanie potencjałów i analiza dźwięku
Potencjały czynnościowe w pojedynczych włóknach nerwowych nerwu słuchowego można wywołać eksperymentalnie. Liczba bodźców zależy od natężenia dźwięku i częstotliwości badanej, każde włókno nerwowe i włókna błony podstawnej są reprezentatywne dla określonej częstotliwości. Przy działaniu dźwięków o wyższych częstotliwościach większa liczba włókien nerwowych ulega pobudzeniu. Liczba pobudzonych włókien nerwowych i wyzwolonych potencjałów czynnościowych koreluje z natężeniem działającego bodźca akustycznego. Tak więc w miejscu dla określonej częstotliwości występuje maksymalne wychylenie błony podstawnej i zgrupowanych tam włókien nerwowych oraz przetworzenie bodźca mechanicznego w impuls nerwowy. Bodziec akustyczny jest następnie dalej przewodzony w ośrodkowej drodze słuchowej. Pierwszy etap selektywnego odbierania dźwięku w odniesieniu do progu percepcji i częstotliwości odbywa się w ślimaku. W drodze słuchowej w pojedynczych neuronach, dzięki mechanizmom transmiterowym, zachodzi różnicowanie tonów i ich natężenia oraz rozumienie mowy opartej na wzorcu rozpoznawania mowy. Słyszenie obuuszne zapewnia możliwość słyszenia kierunku, z którego dźwięk pochodzi. Istotną rolę w określeniu tego kierunku odgrywają różnica faz, czasów w jakich fale docierają od źródła do jednego i drugiego ucha oraz różnice ich ciśnień akustycznych.
Narząd równowagi
Narząd przedsionkowy zajmuje naczelne miejsce w układzie równowagi, a razem z narządem wzroku, czuciem głębokim i powierzchownym jest odpowiedzialny za utrzymywanie równowagi. Umożliwia on orientację przestrzenną poprzez rejestrację wszystkich rodzajów przyśpieszenia (włączając grawitację). Wykształcony u człowieka system poruszania gałek ocznych warunkuje utworzenie ostrego obrazu przedmiotów znajdujących się w naszym otoczeniu. Każdy ruch głowy powoduje kompensacyjne ruchy gałek ocznych – zbaczanie pozwalające na utrzymanie obrazu oglądanego przedmiotu (odruch przedsionkowo-okoruchowy). Na drodze odruchu przedsionkowo-ocznego wywoływany jest rytmiczny ruch gałek ocznych (oczopląs), mający fazę szybką i wolną.
1. Narząd kamyczkowy (statolitowy, otolitowy)
Poziomo ułożona plamka łagiewki (macula utriculi) i pionowo położona plamka woreczka (macula sacculi) i reagują wyłącznie na przyśpieszenie liniowe. Przez stały wpływ przyciągania ziemskiego narząd otolitowy pośredniczy również w odczuwaniu położenia przestrzennego głowy. Grawitacja i dodatkowe przyspieszenia liniowe powodują równoległe przesunięcia kamyczków i odchylenie włosków czuciowych w stosunku do komórek zmysłowych. Bodźcem skutecznym dla komórek zmysłowych ślimaka i przedsionka są siły nożycowe, których wektory działają równolegle do powierzchni komórek. Siły działające prostopadle w stosunku do powierzchni komórek zmysłowych ślimaka i przedsionka nie są bodźcem skutecznym. Po zniesieniu siły grawitacji kamyczki błędnikowe (otolity) przestają spełniać swoją funkcję, ponieważ nie jest możliwe rejestrowanie przyspieszenia liniowego. Przy ruchu pęczka rzęsek w kierunku kinocilium zostaje podwyższony potencjał receptorowy i aktywność spoczynkowa, a przy ruchu w innym kierunku następuje hamowanie aktywności spoczynkowej. Warstwa galaretowata leżąca pomiędzy błoną kamyczkową (otolitową), a nabłonkiem zmysłowym umożliwia tylko ruch równoległy rzęsek. O ułożeniu głowy w stosunku do tułowia informują receptory zlokalizowane w mięśniach i stawach szyi.
2. Układ kanałów półkolistych
Włoski komórek zmysłowych położone są w galaretowatym osklepku przedzielającym światło bańki kanału półkolistego. Wszelkie przesunięcia śródchłonki kanałów półkolistych prowadzą do wychyleń osklepka, a co za tym idzie wychylenia rzęsek komórek zmysłowych grzebieni bankowych. Przesunięcie śródchłonki w przewodzie śródchłonkowym zostało przedstawione w 1873/1874 r. przez Macha i Breuera, a występuje przy ruchu obrotowym głowy. Można je również wywołać stosując bodźce termiczne na przewody półkoliste albo poprzez bodźce mechaniczne (ucisk na błoniasty kanał półkolisty).
Osklepek (cupula) reaguje na przyspieszenie kątowe (obrotowe). Przestrzenne ułożenie kanałów półkolistych zapewnia rejestrację obrotu wokół każdej osi. Ruch śródchłonki przy przyspieszeniu kątowym wywołuje wychylenia osklepka. Wychylenie osklepka z przesunięciem włosków komórek zmysłowych jest bodźcem adekwatnym dla komórek zmysłowych. Wartości progowe bodźca dla kanałów półkolistych poziomych są niższe niż dla kanałów półkolistych pionowych. Bodźcem skutecznym przy pobudzeniu komórek zmysłowych kanałów półkolistych jest również bodziec, którego wektory działają równolegle do powierzchni włosków komórek zmysłowych. Dzięki swojej sztywnej strukturze po wychyleniu osklepek wraca do ułożenia spoczynkowego. Ruchy osklepka mają charakter aperiodyczny. Ruch włosków komórek zmysłowych w kanale półkolistym poziomym w kierunku do łagiewki zwiększa aktywność spoczynkową, a w kierunku od łagiewki zmniejsza aktywność spoczynkową.
3. Przetwarzanie energii i przewodzenie bodźca
Bodźce adekwatne (przesunięcie błony kamyczkowej plamki lub wychylenie osklepka) wywołują pobudzenie komórek zmysłowych (komórki włosowate). Wówczas energia mechaniczna ulega przetworzeniu w energię elektryczną. Miejscowe pobudzenie komórek zmysłowych przekazywane jest do włókien nerwowych. Potencjały czynnościowe nerwów wyzwalane są zgodnie z zasadą „wszystko albo nic”, tak więc częstotliwość impulsów nerwowych zależy od siły bodźca. Bodziec dociera do istoty siatkowatej (formatio reticularis) i pęczka podłużnego tylnego.
Teorie słyszenia
Przemieszczenie podstawy strzemiączka pod wpływem dźwięku powoduje ruch płynów ucha wewnętrznego, który odchyla błonę podstawną ku górze, powodując ugięcie rzęsek komórek słuchowych. Ugięcie rzęsek jest bodźcem, który wyzwala w komórkach słuchowych zewnętrznych procesy biochemiczne powodujące skurcz tych komórek, podobny do skurczu komórek mięśniowych. Skurcz komórek słuchowych zewnętrznych dodatkowo zwiększa wychylenie błony podstawnej i sprawia, że jest ono bardziej precyzyjne, tzn. powstaje w miejscu, w którym znajdują się komórki odpowiedzialne za słyszenie danej częstotliwości. Czynność motoryczna komórek zewnętrznych powoduje wzmożony przepływ płynu w określonej części narządu Cortiego, wywołując ruch rzęsek komórek słuchowych wewnętrznych i w ten sposób ich pobudzenie. Pobudzenie komórek słuchowych wewnętrznych powoduje wydzielanie substancji chemicznych, dzięki którym powstaje bodziec w nerwie słuchowym. W ten sposób bodziec mechaniczny (fala akustyczna) zostaje zamieniony na bodziec elektryczny (zmiana potencjału elektrycznego nerwu słuchowego). Pobudzenie elektryczne jest przez nerw słuchowy przewodzone do wyższych ośrodków nerwowych w korze mózgowej, w której powstaje wrażenie dźwięku. Nerw słuchowy jest uformowany przez neurony zwoju spiralnego ślimaka. W obrębie nerwu słuchowego włókna nerwowe wykazują organizację tonotopową, tzn. włókna odpowiedzialne za przewodzenie informacji o poszczególnych częstotliwościach są zgrupowane razem. Podobna organizacja tonotopowa występuje na wyższych piętrach układu słuchowego w mózgu: jądrze ślimakowym, jądrze górnym oliwki, jądrach wstęgi bocznej, wzgórka dolnego i ciała kolankowatego przyśrodkowego. Integracja obuuszna występuje na dolnym poziomie zespołu jąder oliwki. Lokalizacja bodźców słuchowych występuje na górnym poziomie zespołu jąder oliwki. Ośrodek korowy znajduje się w zakrętach skroniowych poprzecznych. Również w ośrodkach korowych istnieje organizacja tonotopowa.
Teorie słyszenia
Teoria rezonansowa
Teoria fali stojącej
Teoria fali biegnącej
Teorie słyszenia stanowiły i stanowią dotychczas przedmiot dociekań fizjologów i akustyków. Chodzi tu o sposób, w jaki drgania bębenka, przeniesione za pośrednictwem układu kostek ucha środkowego do ucha wewnętrznego, powodują pobudzenie określonych zakończeń nerwowych w organie Cortiego.
Kluczowym zagadnieniem jest sposób przeniesienia drgań strzemiączka na błonę podstawową, z którą jest związany organ Cortiego. Istnieją tu trzy możliwości odpowiadające różnym ujęciom teoretycznym:
Teoria rezonansowa – zakłada się, że ze względu na małe rozmiary ślimaka można uważać, że zaburzenia wytworzone wskutek drgań strzemiączka rozchodzą się natychmiast w całej objętości ślimaka. Gdy strzemiączko drga z bardzo małą częstotliwością, wytworzone przezeń zmiany ciśnienia powodują przepływ perylimfy w obu kierunkach przez dziurkę osklepka ślimaka i wymuszenie drgań błonki zamykającej okienko ślimaka. Gdy częstotliwość drgań jest znaczna, perylimfa nie zdąży przepływać przez dziurkę osklepka i aby wyrównać ciśnienia między schodami przedsionka i bębenka, przewód ślimakowy a wraz z nim błona podstawowa musi choć w jednym miejscu drgać, przesuwając nadmiar cieczy. Ponieważ podatność błony podstawowej zmienia się nieznacznie wzdłuż przewodu ślimakowego, przeto przy większych częstotliwościach muszą drgać części błony podstawowej leżące bliżej przedsionka itd.
Przeciwko teorii rezonansowej przemawiają zastrzeżenia wynikające ze zdolności ucha do rozróżniania tonów o nieznacznie różniących się częstotliwościach i szybko następujących po sobie.
Teoria fali stojącej – wyodrębnienie częstotliwości następuje w wyniku właściwego dla każdej częstotliwości rozkładu fal stojących wzdłuż błony podstawowej, a więc i przestrzennego rozkładu pobudzania tej błony. Wynikałoby stąd, że przeprowadzaniem analizy dźwięków jest obarczony wyłącznie mózg.
Ze względu na to, że rozkład fal stojących, wywołanych dźwiękiem złożonym, zależy od wzajemnych faz tonów składowych, ucho powinno odczuwać różnice faz, czego doświadczalnie nie stwierdzono.
Teoria fali biegnącej – głoszona przez znakomitego uczonego G. Bekesy. Stwierdził on, że pod wpływem drgań strzemiączka powstają w błonie podstawowej fale, rozchodzące się od przedsionka ku dziurce osklepka. Amplitudy tych fal w miarę oddalania się od przedsionka wolno wzrastają, osiągając w pewnym miejscu błony podstawowej wartość największą, a następnie szybko zanikają. Miejsce występowania największej amplitudy drgań zależy od częstotliwości i dla największych słyszalnych częstotliwości występuje w części błony najbliższej przedsionka, dla najmniejszych zaś – w części najbliższej dziurki osklepka.
Dodatkowe zaostrzenie właściwości selektywnych jest związane z właściwościami układu nerwowego.
Granice i powierzchnia słyszalności
Zakres częstotliwości słyszalnych jest ograniczony budową anatomiczną ucha. Najmniejsza częstotliwość słyszalna wynosi przeciętnie 16Hz, największa zaś 20kHz, jednak ta częstotliwość obniża się z wiekiem do 16kHz, a nawet do 10kHz.
Podobnie istnieją granice związane z natężeniem dźwięku. Wartości ciśnień akustycznych dźwięków, przy których ucho zaczyna odbierać wrażenia dźwiękowe, i ciśnień wywołujących wrażenie bólu, określające granicę słyszalności, zostały zmierzone przez wielu badaczy. Wyniki średnie obrazuje poniższy wykres:
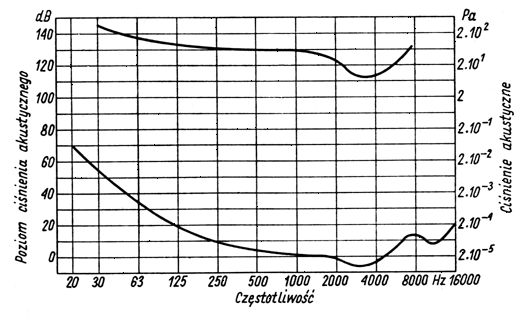
Granice słyszalności ludzkiego ucha
Krzywa dolna, odpowiadająca najmniejszym akustycznym ciśnieniom słyszalnym, nazywa się dolną granicą słyszalności. Górna krzywa natomiast, odpowiadająca ciśnieniom, przy których zaczyna występować uczucie bólu w uszach, nazywa się granicą słyszenia bolesnego lub ogólnie – górna granicą słyszalności
Przedłużając dolną i górną granicę w obie strony aż do przecięcia otrzymuje się wewnątrz nich powierzchnię, zwaną powierzchnią słyszalności.
Wysokość i natężenie dźwięku
Wysokość tonu
Poziom głośności
Głośność
Wysokość tonu jest wrażeniem słuchowym, umożliwiającym określenie położenia tonu na skali częstotliwości, a za jego jednostkę przyjęto mel. Wrażenie wysokości tonu zależy od natężenia dźwięku. Na podstawie pomiarów ustalono związek między subiektywna skalą wysokości tonu a obiektywną skala częstotliwości (przy założeniu, że wysokość tonu o częstotliwości 1000Hz i poziomie ciśnienia akustycznego równym 40dB jest 1000 melów):
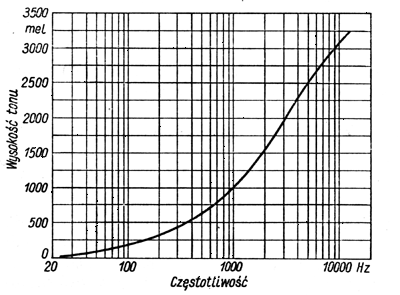
Wrażeniem subiektywnym związanym z natężeniem dźwięku jest głośność, zależna również od częstotliwości. Jednostką poziomu głośności jest fon, któremu odpowiada poziom natężenia dźwięku tonu odniesienia o częstotliwości 1000Hz.
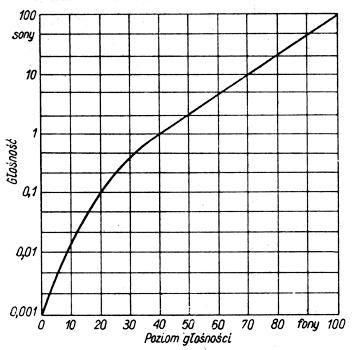
Jednostką głośności natomiast jest son, który odpowiada głośności tonu o poziomie głośności 40 fonów. Zależność między głośnością tonów wyrażoną w sonach a poziomem głośności w fonach przedstawia poniższy wykres:
Ucho wewnętrzne może być pobudzane przez drgania błony bębenkowej i kosteczek słuchowych i mówimy wtedy o przewodnictwie powietrznym. Możliwe jest też pobudzenie ślimaka bez udziału ucha środkowego i wtedy mówimy o przewodnictwie kostnym. Ogólnie ten rodzaj przewodzenia dźwięku polega na pobudzeniu ślimaka poprzez drgania kości czaszki. Jest to mniej efektywny sposób przekazywania energii akustycznej do ucha wewnętrznego, gdyż czaszkę dużo trudniej wzbudzić do drgań niż błonę bębenkową. Jednak badanie przewodnictwa kostnego jest bardzo istotne, gdyż dostarcza cennych informacji o funkcjonowaniu ucha środkowego i zewnętrznego.
Wyobraźmy sobie, że pacjent ma np. zatkany woskowiną przewód słuchowy zewnętrzny. Oznacza to, że błona bębenkowa jest zasłonięta i nie będzie mogła drga tak silnie. W rezultacie pacjent słyszy gorzej (na drodze powietrznej). Jeżeli teraz ominiemy tę uszkodzoną część narządu słuchu i wzbudzimy ślimak drogą kostną, np. przykładając drgający stroik (kamerton) do czaszki, to okaże się, że pacjent słyszy ton dobrze, jakby nie miał ubytku. Oznacza to, że ślimak działa poprawnie, tylko nie jest wystarczająco pobudzany na drodze powietrznej. Wniosek: uszkodzenie znajduje się gdzieś w uchu środkowym lub, jak w naszym przypadku, w uchu zewnętrznym. Jeżeli teraz zbadamy innego pacjenta, u którego okaże się, że tak samo źle słyszy na drodze kostnej i powietrznej, to znaczy, że niestety uszkodzone jest ucho wewnętrzne – nie ma znaczenia, jak pobudzimy ślimak – i tak słyszy on źle.
Pytanie: Czy będzie jakaś różnica między przewodnictwem kostnym a powietrznym, gdy uszkodzony będzie np. i ślimak, i ucho środkowe?
Tak, ponieważ badając słuch na drodze powietrznej, na stopień niedosłuchu będzie miało wpływ i uszkodzone ucho środkowe, i ślimak, natomiast w przypadku przewodnictwa kostnego pominięte jest ucho środkowe i o stopniu ubytku słuchu decyduje jedynie ślimak.
Umiejscawianie źródła dźwięku
Aby prawidłowo określić położenie źródła dźwięku, trzeba prawidłowo wyznaczyć współrzędne biegunowe tego obiektu: długość promienia, czyli odległość głowy słuchającego od źródła, azymut, czyli kąt między promieniem a płaszczyzną symetrii głowy, i zenit, czyli kąt między promieniem a płaszczyzną poziomą przechodzącą przez środki uszu.
Najłatwiejsza jest ocena azymutu, najtrudniejsza – zenitu.
Lokalizacja źródła dźwięku dla:
azymutu: w zakresie małych częstotliwości (poniżej 800Hz) określanie kierunku źródła odbywa się na podstawie różnicy faz (a właściwie opóźnień) między falami dźwiękowymi dobiegającymi do obu uszu w zakresie dużych częstotliwości – na podstawie różnicy ciśnień akustycznych lokalizacja źródła dźwięku na podstawie różnicy barwy dźwięku (ucho odwrócone od źródła słabiej słyszy dźwięki o dużych częstotliwościach)
zenitu: zasadniczą rolę odgrywają tu ruchy głowy odległość źródła dźwięku – w przestrzeni otwartej ocena opiera się przede wszystkim na odczuwaniu zmiany barwy dźwięku (im większa częstotliwość dźwięku tym większe pochłanianie energii akustycznej w atmosferze) oraz jego głośności, stąd konieczna jest uprzednia znajomość źródła zarówno co do barwy dźwięku, jak i natężenia.
AUDIOMED APARATY SŁUCHOWE
ul. M. Reja 16/18 lok. 2
87-800 Włocławek
tel: 54 426 25 85, 604 479 703